
Na primeira parte desta série, discuti o lado esquecido da água e a imensa complexidade que as moléculas de água podem demonstrar como um grupo. Infelizmente, embora existam amplas evidências de que há mais água do que imaginamos, a comunidade científica se recusou firmemente a reconhecer esse fenômeno, e como esse negacionismo generalizado espelha muitos dos problemas que enfrentamos ao longo da pandemia. Eu aconselho a leitura primeiro, pois ele estabelece o contexto para as consequências biológicas da água EZ (água cristalina líquida) que será discutido aqui.
Introdução
Gerald Pollack baseou-se neste século de observações, e eventualmente determinou que, quando uma superfície carregada negativamente está presente, e a energia eletromagnética ambiente, como a luz, existe (até certo ponto, o som também pode cumprir esse papel), a água se reunirá em camadas de folhas hexagonais offset com a fórmula H3O2 .
Esta estrutura tem um grau significativo de solidez, e expulsará a maioria das coisas de entrar nela (por exemplo, microesferas de poliestireno), incluindo os átomos de hidrogênio deslocados (como é H 1,5 O e não H2O).
Esses átomos H deslocados carregados positivamente (doravante chamados de prótons), por sua vez, se reúnem imediatamente fora dessa rede, criando assim um gradiente de pH e carga que pode ser medido.
Em muitos casos, essa estrutura H3O2 pode ser enorme – em condições favoráveis, Pollack e outros mediram aquelas que variam de 0,1 milímetros a 0,5 milímetros (100.000 a 500.000 nanômetros ou nm) de tamanho. As células dependem dessa água, por isso contêm um grande número de superfícies a partir das quais a água pode se formar.
Por exemplo, 20% da célula é ocupada por seu citoesqueleto (uma rede de proteínas que mantém sua estrutura), e a análise de micrografias eletrônicas de alta voltagem mostrou que, dentro do citoesqueleto, mais da metade da água presente está a 5 nm de uma superfície que poderia potencialmente formar H 3 O 2 (nota: uma molécula de H 2O tem 0,27 nm de tamanho).
Uma vez que os sítios superficiais estruturantes da água (H 2 O) estão tão intimamente agrupados nas células, é compreensível por que a água líquida cristalina (EZ, H3O2) seria tão mais aparente para os indivíduos que observam as células do que para os trabalhadores que realizam trabalho não celular observando a água ao microscópio.
Isso, por sua vez, levanta uma questão diferente: por que as células são projetadas para criar tanta água líquida cristalina (H3O2)?
Mistérios da Célula
No artigo anterior, discuti uma questão comum que observo dentro da ciência. Quando um modelo incorreto é utilizado para explicar um processo natural, discrepâncias entre o modelo e a realidade inevitavelmente aparecerão.
Seria de esperar que, quando isso acontecesse, encorajaria aqueles que defendem o modelo errôneo a reexaminar seu modelo, mas, em vez disso, como tanto foi investido nesse modelo, eles denunciarão quaisquer desafios a ele e criarão inúmeras maneiras criativas de explicar cada falha do modelo existente.
Da mesma forma, com a biologia celular, nosso conhecimento da célula é surpreendentemente primitivo e os modelos existentes muitas vezes não conseguem explicar o que ocorre dentro do ambiente celular.
No entanto, como modelos melhores não estão ao alcance da comunidade científica, fomos forçados a remendar continuamente os modelos existentes para que eles possam dar conta dos inúmeros mistérios da vida.
Neste ponto, acredito que uma das principais causas dessa situação é a pesquisa científica se distorcer para priorizar o foco em descobertas com as quais a indústria pode lucrar.
Por exemplo, a imunologia tem um foco estreito nos aspectos do sistema imunológico, que podem ser alvo de vacinação ou medicamentos patenteados, e porque isso deixou muitos outros componentes da resposta imune negligenciados, levando a ser comumente referido como um dos sistemas menos compreendidos no corpo.
Da mesma forma, como as drogas farmacêuticas geralmente funcionam afetando receptores e canais nas células, a biologia celular adotou um foco tacanho nesses aspectos de uma célula.
Felizmente, a fase líquida cristalina (EZ) da água fornece uma variedade de explicações para muitos dos fenômenos que os modelos existentes não abordam adequadamente. Neste artigo, vou me concentrar em alguns deles.
Nota: Muito do que é discutido neste artigo é abordado em mais detalhes dentro desses três livros (da mesma forma, a maioria das referências para este artigo são provenientes desses livros, então não vou citá-las repetidamente ao longo do artigo).
Se você quiser estudar mais sobre o assunto, recomendo a leitura desses livros (todos de autoria de Gerald H. Pollack) nesta ordem:
A Quarta Fase da Água: Além do Sólido, do Líquido e do Vapor (2013)•Células, Géis e os Motores da Vida: Uma Nova Abordagem Unificante da Função Celular (2001)•Transições de Fase em Biologia Celular (2008) – esta é a mais técnica das três.
Integridade Celular
Um dos grandes quebra-cabeças da biologia é a imensa durabilidade que as células têm. Se você considerar o modelo clássico – células sendo sacos de líquido contidos dentro de uma membrana de mosaico de fluido, deve ser fácil para forças externas “estourar” células e ter todo o seu conteúdo derramado.
No entanto, na maioria dos casos, as células mantêm sua integridade apesar de estressores significativos. As células podem sobreviver a traumas, incluindo serem guilhotinadas ao meio, desenhadas e esquartejadas (para que componentes específicos possam ser isolados e trabalhados – como ao realizar (fertilização in vitro), ou atiradas cheias de buracos com balas elétricas, cada uma das quais seria suficiente para “estourá-las”.
No entanto, em cada caso, a integridade celular do componente restante persiste. Da mesma forma, se a membrana de uma célula é removida, seu conteúdo interno permanece no lugar em vez de desaparecer rapidamente.
Também é sabido há mais de 50 anos que, se as fibras musculares perdem suas membranas, sua capacidade funcional (criando uma força contrátil) permanece intacta.
Três pistas ajudam a explicar esses fenômenos:
. A primeira é que, como Pollack mostrou, as gotículas de água têm uma quantidade notável de integridade e, como as células, manterão sua integridade (e posteriormente se fundirão novamente) se uma microlâmina for usada para guilhotiná-las ao meio.
. A segunda é que Pollack também mostrou que as gotículas de água contêm um grau significativo de água líquida cristalina, o que provavelmente é o que confere sua integridade.
. A terceira é que as moléculas de água nas células existem predominantemente dentro de géis, que são compostos dessa mesma água líquida cristalina.
Dito de outra forma, isso significa que a estabilidade de uma célula é em grande parte uma propriedade de sua água que a mantém unida, em vez da estrutura externa que a encapsula. Outro aspecto importante da arquitetura celular deve agora ser considerado. Como as condições para a formação de água líquida cristalina estão presentes em toda a célula (superfícies hidrofílicas carregadas negativamente e energia infravermelha ambiente), as células devem rapidamente estar se enchendo com camadas de água cristalina líquida de centenas de micrômetros de espessura.
No entanto, ao longo das células, as superfícies geralmente estão apenas frações de um micrômetro de distância. Isso significa que a estrutura da célula depende do crescimento de água cristalina líquida, mas simultaneamente restringe essa estrutura cristalina de crescer até seu tamanho total.
Biotensegridade
Classicamente na arquitetura, os edifícios são criados por terem esqueletos fortes sobre os quais o resto da estrutura se apoia. Por exemplo, antigamente, usávamos frequentemente pilares de pedra; hoje em dia, os arranha-céus necessitam de superestruturas de aço para atender às necessidades destes edifícios:

Este modelo muitas vezes não funciona dentro de organismos vivos, porque a vida, ao contrário desses edifícios, requer movimento rápido, e os organismos simplesmente não podem produzir estruturas sólidas com a mesma resistência que as vigas de aço.
No entanto, um modelo arquitetônico alternativo e mais complexo tem sido desenvolvido, o qual é frequentemente utilizado por aqueles que abraçam a complexidade.
Tensegrity (abreviação de integridade tensional), foi um modelo proposto pela primeira vez por Buckminster Fuller. Ele postula que se uma série de estruturas não-compressíveis são ligadas entre si por uma rede de conexões elásticas (que podem armazenar tensão quando esticadas), uma estrutura muito mais forte é criada.
Isso ocorre porque qualquer força que a estrutura recebe será igualmente distribuída através de cada uma dessas conexões elásticas em vez de se concentrar em um único componente (por exemplo, o pilar de pedra) e, portanto, muito menos provável de exceder o ponto de ruptura de qualquer componente estrutural único.
O trabalho de Fuller posteriormente inspirou numerosos edifícios a serem construídos sobre os princípios da tensão. Esta é uma imagem clássica dele segurando uma esfera tensa que ele fez:

A biotensegridade encapsula a constatação de que esse mesmo sistema também ocorre em toda a natureza. Para o corpo humano, em cada nível de organização, está presente uma matriz de tração ligada que lhe confere estabilidade.
Um cirurgião de mão francês, Jean-Claude Guimberteau, também fez um trabalho notável ao demonstrar a presença de estruturas de tração ligadas ao nível da fáscia (um tecido conjuntivo presente em todo o corpo com o qual muitos terapeutas manuais trabalham), através de pequenas câmeras (ampliadas) colocadas no corpo durante cirurgias minimamente invasivas:
Com o passar dos anos, parece haver um maior consenso dentro do campo médico integrativo de que a biotensegridade é um modelo válido para a compreensão do corpo, e que redes ligadas de tensão estão presentes do maior para o menor nível do corpo (por exemplo, o citoesqueleto é a unidade de conexão elástica dentro de cada célula).
No entanto, embora essa teoria seja frequentemente discutida, ainda há duas grandes questões não abordadas com ela, que acredito que a água cristalina líquida pode explicar. A primeira é que, no trabalho de Guimberteau, ele observou que minúsculos vacúolos (compartimentos fechados cheios de água) por todo o corpo formam a unidade básica incompressível da qual grande parte da tensão do corpo depende.
A segunda é que, em uma ampliação ainda maior dentro das células (que também não possuem uma unidade incompressível claramente definida), enquanto um citoesqueleto elástico está presente, nada o mantém visivelmente sob tensão (é altamente discutível se as conexões da célula com a matriz extracelular são suficientes para fazer isso).
Isso é importante porque a resistência estrutural da tensegridade só emerge quando os componentes de uma estrutura estão sob tensão. No caso do primeiro, acredito que os microvacúolos preenchidos com água líquida cristalina (EZ) (criando assim sua incompressibilidade).
No caso das gotículas de água, Pollack concluiu que a repulsão mútua entre prótons carregados positivamente (empurrados para o centro da gota por zonas de exclusão) cria uma força externa resistida pelo limite cristalino líquido da gota. O equilíbrio entre essas duas forças faz com que a gotícula assuma uma forma esférica, e acredito que o mesmo mecanismo esteja em ação nos microvacúolos em todo o corpo humano.
No caso deste último, é importante lembrar quanta expansão é produzida por proteínas que criam géis (muitos géis têm mais de 99,9% de água). Pollack, por sua vez, mostrou que a água cristalina líquida que está constantemente tentando se formar dentro da célula é incapaz de atingir seu tamanho total devido às proteínas resistirem ao estiramento que teria que ocorrer para que o gel cristalino líquido (que as proteínas circundantes geram) pudesse atingir seu tamanho máximo.
Isso acontece tanto no nível da célula (como o citoesqueleto se estende à medida que a célula se expande até seu tamanho máximo, até que não possa permitir que ocorra mais expansão) e dentro de proteínas por toda a célula. No nível de proteína, o corpo muitas vezes depende de ligações químicas a serem feitas entre proteínas para restringir a expansão máxima que pode ocorrer em resposta a uma formação de gel.
Além disso, as proteínas podem alternar entre uma conformação dobrada e uma desdobrada (por exemplo, uma hélice versus uma bobina), algo frequentemente determinado pela tensão aplicada à proteína (como a pressão expansiva da água cristalina líquida dentro de um gel). Há uma variedade de consequências importantes desta mudança de conformação que serão discutidas mais adiante com a fisiologia dos músculos.
Além disso, muitos outros componentes do corpo também parecem depender de água cristalina líquida:Estudando o colágeno, Melacini e col. observaram a importância da água na estabilização de sua estrutura cristalina de tripla hélice.
Eles descobriram que a água dentro da hélice de colágeno forma uma “estrutura semiclatrato (Um Clatrato é um composto de inclusão, no qual moléculas de uma substância são confinadas em cavidades formadas pela molécula hospedeira[1], originando um agregado supramolecular), que envolve e interconecta triplas hélices na rede cristalina”.
A água dentro da matriz óssea também é altamente estruturada, uma estrutura que tem sido encontrada associada não apenas a macromoléculas orgânicas, como colágeno e proteoglicanos, mas também às superfícies minerais.
A água, de fato, parece desempenhar um papel fundamental na orientação de nanopartículas minerais em arranjos paralelos dentro da matriz óssea, fornecendo essa orientação mesmo na ausência de moléculas orgânicas.
Expansões de Gel para o Dia a Dia
Muitas tecnologias com as quais estamos familiarizados (por exemplo, fraldas) dependem de hidrogéis que podem se expandir em uma estrutura semissólida que retém água. Da mesma forma, podemos observar diretamente que muitos processos maiores dentro do corpo também dependem da tendência da água de se agrupar nas estruturas cristalinas líquidas maiores.
Como a arquitetura normal do comportamento de reticulação da proteína limita a quantidade de géis que podem se expandir (já que as proteínas que precisariam se separar para acomodar o gel em crescimento são impedidas de fazê-lo pelas ligações cruzadas), quando o tecido ou a proteína é danificado, essa limitação pode ser parcialmente removida.
Como resultado, experimentos em escala microscópica foram realizados mostrando que o tecido humano incha e se expande quando é danificado. Da mesma forma, Pollack argumentou que isso é provavelmente o que acontece quando você sofre um trauma musculoesquelético.
No modelo proposto por Pollack, a formação inicial do gel expande ainda mais a lágrima existente, e essa expansão progressiva da água líquida cristalina eventualmente leva ao inchaço visível.
Em uma escala maior, um dos maiores desafios de engenharia para o corpo é fazer com que as articulações de sustentação de peso, como os joelhos, sejam capazes de manter sua amplitude de movimento sem serem danificadas pelo atrito contínuo que experimentam diariamente.
Uma característica notável sobre a água cristalina líquida é que, desde que a superfície a partir da qual ela se forma permaneça, ela pode ser destruída e, em seguida, reformar sua superfície quase sem atrito repetidamente.
Por conta disso, a água líquida cristalina acaba sendo o componente que absorve o estresse experimentado por articulações saudáveis, e desde que a articulação esteja saudável, essa água pode se regenerar instantaneamente a partir desse estresse.
Em conjunto com esta camada regeneradora de água cristalina líquida carregada negativamente, no centro da articulação, há uma bolsa de prótons carregados positivamente que se repelem uns dos outros e criam uma pressão expansiva (como o que é visto em uma gota de água).
Como a cápsula articular sela essa região, esses prótons são incapazes de escapar e, assim, funcionam efetivamente como ímãs repelentes (por exemplo, considere um trem maglev) que resistem ao peso do corpo e mantêm o espaço central dentro da articulação.
Uma das coisas que considero mais atraentes sobre o modelo de Pollack para as articulações é a qualidade específica do líquido sinovial dentro da articulação do joelho.
Quando você o vê em uma câmera durante uma cirurgia artroscópica, a difusão dentro dele é visivelmente lenta, enquanto se você extraí-lo diretamente (por exemplo, durante uma aspiração do joelho), você pode dizer que ele tem uma qualidade muito mais espessa e gelatinosa, outra qualidade que aprendi a associar à presença de água cristalina líquida.
Pollack também argumenta que seu modelo sugere que o comportamento da água EZ (líquido cristalino) se assemelha ao de uma clara de ovo gelatinosa que é semissólida quando deixada sozinha, mas capaz de fluir em resposta a uma força de cisalhamento imposta.
Gradientes Celulares
Nota: um “gradiente” descreve uma diferença na concentração de coisas em duas áreas diferentes. Isso pode incluir cargas elétricas (as baterias dependem de gradientes), eletrólitos ou temperaturas.
O paradigma existente da física suporta o seguinte:
- O estado natural das coisas deve ser desordenado e uniformemente misturado.
- Sempre que você faz algo se tornar mais ordenado (por exemplo, formando um cristal ou criando um gradiente entre duas áreas), a energia deve ser gasta para criar essa ordem.
- Quando uma estrutura ordenada se torna desordenada, a energia é liberada no processo que às vezes pode ser colhida (por exemplo, queimando lenha em um fogão a lenha para aquecer nossas casas).
- Sempre que a energia é liberada de algo, se você tentar capturar e armazenar essa energia, alguma energia sempre será perdida.
Essas leis, por sua vez, são usadas para refutar a possibilidade de que qualquer tipo de sistema do tipo “energia livre” possa existir, como o sistema de água de zona de exclusão (EZ) proposto por Pollack.
Infelizmente para o paradigma existente, a biologia muitas vezes parece violar essas leis, pois está continuamente se movendo em direção a um estado mais ordenado em vez do estado desordenado que o paradigma prevê.
A resolução atual para esse paradoxo (que ganhou um prêmio Nobel) é que os organismos vivos funcionam como “estruturas dissipativas”, que trocam a ordem em grandes quantidades de componentes ordenados que acumulam de seu ambiente em troca de dar ordem a seus próprios conteúdos desordenados.
Embora até certo ponto isso permita que o paradigma existente se sustente, não acredito que seja totalmente preciso, pois a água, através de sua estrutura cristalina líquida, tem a capacidade de armazenar energia radiante que passa por ela e converter essa energia para ordem que o corpo pode utilizar.
O gradiente de sódio e potássio
Uma das coisas que as células vivas são bem conhecidas por fazer é concentrar potássio dentro de si mesmas e expulsar reciprocamente o sódio. Como a concentração dentro e fora das células é diferente, existe um gradiente, que pelas leis existentes da física deveria tentar se igualar e desaparecer rapidamente.
Como isso não acontece, o modelo existente argumentou que a membrana celular impede a passagem da maioria, mas não de todo o sódio e potássio (inibindo assim o gradiente de se equalizar) e que existem bombas de sódio e potássio na membrana celular que trocam continuamente o sódio dentro da célula pelo potássio fora da célula.
Como essa troca é tão vital para manter a saúde da célula, um grande foco na biologia celular é colocado na importância da bomba de troca de sódio e potássio.
Infelizmente, há três problemas fundamentais com esse modelo (a evidência de cada um deles e mais é apresentada por Gilbert Ling aqui):
- A matemática não se soma – as bombas de sódio e potássio existentes simplesmente não têm a capacidade de neutralizar o desfazimento natural dos gradientes de sódio e potássio. Por exemplo, em células musculares, manter o gradiente de sódio e potássio com bombas requer entre 4 a 30 vezes a energia total disponível na célula.
- As células são capazes de manter um gradiente com uma variedade de outros componentes indesejáveis que expelem (por exemplo, bactérias expelindo antibióticos) e, para explicar esses fenômenos, mais e mais bombas são identificadas para tentar apoiar o modelo. Isso é um problema porque é improvável que as células tenham a capacidade de sustentar simultaneamente tantas bombas diferentes.
- Ling pegou células musculares de rã cujo ambiente foi alterado para que ficassem completamente sem energia (que é necessária para operar a bomba de sódio e potássio).
Apesar disso, o gradiente foi mantido. Isso então levanta a questão do que poderia estar criando esse gradiente? Ling fez as seguintes observações:
- O gradiente foi mantido se a membrana celular fosse removida.
- A produção de uma membrana com bombas de sódio e potássio que não continham um citoplasma (o interior de uma célula) resultou no desaparecimento rápido do gradiente do interior da membrana. Isso sugere que o gradiente é uma propriedade do citoplasma celular e não da membrana celular. Curiosamente, a propriedade idêntica foi encontrada em géis artificiais, que assim como o citoplasma, também geram grandes quantidades de água líquida cristalina:
Mostramos que a barreira de gel é capaz de manter uma separação estável de soluções iônicas de diferentes forças iônicas e composições químicas sem qualquer atividade de bombeamento. Para o gradiente de concentração de Na+ /K+ sustentado através da barreira, um potencial elétrico negativo se desenvolve dentro do lado rico em K+.
A situação lembra a da cela. Além disso, também o fluxo advindo de moléculas de água através da barreira do gel é restrito, apesar dos poros grandes do gel e dos gradientes de pressão osmóticos ou hidrostáticos através dele. Esse fenômeno de concentração também foi observado por outros autores:
Evidências dessa condensação acumulativa foram produzidas há quase um século, quando Bungenberg de Jong, em 1932, mostrou que mesmo soluções diluídas de polímeros, quando agitadas, coalesciam em gotículas – então chamadas de coascervados – nas quais a matéria orgânica se tornava altamente concentrada.
A concentração de polímero em tais gotículas pode exceder a concentração no banho circundante em até 10.000 vezes. Quando colocadas em certos corantes, as gotículas tornaram-se progressivamente mais coloridas, a intensidade muitas vezes superior à da solução circundante. Assim, as gotículas de gel tinham a capacidade de concentrar determinados solutos, uma vez que a célula concentra potássio.
Uma vez que os géis demonstram independentemente muitas das características inexplicáveis das células, é razoável concluir que eles podem compartilhar um mecanismo para produzir o gradiente de sódio e potássio observado ao longo da biologia.
Barreiras
Um grande desafio para todas as células é manter coisas indesejadas fora do sistema de suas células. Classicamente, vemos o processo de entrada de substâncias patogênicas nas células como sendo um produto de sua interação com receptores correspondentes nas células (uma visão de mundo que acredito ser enfatizada pela ciência médica porque drogas podem ser desenvolvidas para interromper esse processo).
Pessoalmente, acredito que esse processo de exclusão pela célula é fortemente influenciado por cargas, potencial zeta e água líquida cristalina. Como a água cristalina líquida só pode se formar em superfícies carregadas negativamente (da mesma forma que as moléculas de água expelem meio átomo de hidrogênio e, assim, mantêm uma carga negativa líquida devido aos hidrogênios perdidos), quase todas as superfícies do corpo (e muitos outros sistemas naturais) são revestidas com carga negativa.
Uma vez que a estabilidade coloidal depende em parte da repulsão mútua criada por cargas idênticas, a maioria dos sistemas coloidais naturais requer cargas negativas para permanecerem dispersas e se amontoarem se cargas positivas excessivas estiverem presentes (a maioria dos métodos de restauração do potencial zeta para ajudar a dispersão coloidal opera com base neste princípio).
Organismos patogênicos muitas vezes transmitem cargas positivas ao seu ambiente (como através de bactérias descarboxilando as cargas negativas de aminoácidos dentro das proteínas). Isso faz com que proteínas e células dentro do corpo comecem a se aglomerar (o parasita da malária é particularmente bom em fazer isso), o que impede que o sistema imunológico tenha acesso aos micróbios e aumenta sua capacidade de obter acesso às células.
Uma das observações particularmente interessantes feitas pelos proponentes originais do potencial zeta fisiológico foi que as infecções bacterianas e virais (acredito que esta foi a mais estudada com influenza) criariam uma degradação consistente do potencial zeta fisiológico.
Isso fornece uma explicação para por que vírus como o influenza podem ser tão mais perigosos para os idosos. Alguém com um potencial zeta saudável pode tolerar um pequeno comprometimento dele (eles apenas se sentirão um pouco indispostos), mas para alguém que já tem um potencial zeta prejudicado (que é uma consequência muito comum do envelhecimento), isso pode ser suficiente para passar um limiar crítico onde a doença grave se inicia.
Uma das primeiras características distintivas da COVID-19 para mim foi que muitos pacientes desenvolveram sinais clínicos sugerindo interrupção potencial zeta grave, algo que considerei altamente incomum.
Uma das principais razões pelas quais eu estava preocupado com a proteína spike muito antes das injeções entrarem no mercado, foi porque eu a identifiquei como a provável culpada pela potencial interrupção do zeta. Isto deveu-se ao facto de ser um componente novo do vírus SARS-CoV-2 (que diferia do SARS-CoV-1) que estava presente na superfície do vírus e que tinha uma elevada densidade de carga positiva.
Desde então, foi demonstrado que a proteína spike induz diretamente a aglomeração de glóbulos vermelhos, provavelmente devido à sua distribuição de carga ter o potencial de criar uma interrupção significativa do potencial zeta fisiológico.
Um dos comentários interessantes que tenho visto repetidamente declarados tanto por indivíduos que tomaram protocolos para melhorar a produção de água cristalina líquida de seu corpo quanto por indivíduos que fizeram o mesmo para melhorar seu potencial zeta, é que eles raramente ficam doentes.
Isso pode potencialmente significar que os sintomas de infecções são muito menores se esses parâmetros fisiológicos já estiverem em boa saúde (por exemplo, uma vez que você não passa de um limite crítico de potencial zeta), ou porque eles impedem diretamente que as células sejam infectadas por patógenos.
Como muitos fatores que afetam positivamente um dos parâmetros geralmente afetam o outro (por exemplo, o aterramento melhora o potencial zeta e promove o crescimento de água cristalina líquida no corpo), é um pouco difícil dizer qual efeito predomina, e uma grande quantidade de pesquisas para esta série foi baseada em tentar responder a essa pergunta.
Para este artigo, olhamos para ele apenas de uma perspectiva de barreira e, nesse ponto, há algumas evidências de corroboração interessantes. O mais antigo que encontrei foi do trabalho de Viktor Schauberger (1885-1958), que acreditava ser extremamente importante que a água viajasse em um padrão de vórtice, algo como discutido na parte 1, demonstrou aumentar a presença de água líquida cristalina.
Uma das muitas descobertas feitas pela utilização de sistemas de vórtice baseados em seu projeto para abastecimento de água foi que ele reduziu o crescimento de bactérias dentro do sistema de água (por exemplo, dentro de tubulações).
Outras pesquisas descobriram diretamente que a presença de água líquida cristalina impede o crescimento e a invasão de bactérias, como este estudo:Em um teste realizado com E. coli, as células penetraram progressivamente EZ [água cristalina líquida] ao longo de 2 dias.
Além disso, o Nafion portador de EZ teve 80% menos acúmulo de biomassa de E. coli ao longo de 2 dias em comparação com uma superfície de óxido de alumínio lisa, hidrofílica e sem EZ. Isso sugere que a EZ pode representar a primeira linha de defesa, espacial e temporalmente, contra bactérias que se aproximam de determinadas superfícies hidrofílicas.
Esses resultados podem ter implicações importantes no desenvolvimento de superfícies de materiais resistentes à bioincrustação para aplicações sensíveis à fixação bacteriana e à formação de biofilme.
E este estudo:
A sericina da seda é uma proteína globular cuja resistência contra incrustações é importante para aplicações em biomateriais e membranas de purificação de água. Aqui é mostrado como a sericina gera uma zona de exclusão de água [líquido cristalino] que pode facilitar o comportamento anti-incrustante.
Microesferas carregadas negativamente foram usadas para mimetizar a carga superficial e domínios hidrofóbicos em bactérias. Imersa em água, a sericina formou uma zona de exclusão de 100 μm (para foulantes de tamanho micron), juntamente com um gradiente de prótons com uma diminuição de >2 unidades de pH.
Assim, quando em contato com a sericina, as moléculas de água próximas à superfície se reestruturam para formar uma barreira física de exclusão que pode impedir a bioincrustação.
A diminuição do pH torna o meio aquoso inviável para bactérias neutrofílicas. Portanto, a resistência à bioincrustação parece explicável, entre outros fatores, com base em fenômenos de exclusão hídrica.
Muitos tecidos dependem da presença de água líquida cristalina que os reveste, e isso é provável, pelo menos em parte, devido à barreira protetora que eles criam.
Green e Otori, em 1970, por exemplo, encontraram zonas de exclusão de aproximadamente 350 μm de profundidade que se estendiam da córnea e também da lente de contato (é importante proteger o olho de danos, e a água líquida cristalina funciona como uma barreira transparente e bem lubrificada, qualidades ambas críticas para os olhos).
Pollack, há aproximadamente 15 anos, também encontrou resultados idênticos ao polyNIPAM, o gel usado para fabricar lentes de contato. Um dos tecidos mais importantes do corpo para fornecer uma camada protetora (mas bem lubrificada) está no revestimento endotelial dos vasos sanguíneos. Lá, grandes quantidades de sangue criando forças de cisalhamento significativas passam constantemente sobre os vasos sanguíneos.
Até este ponto, Malcom Kendrick observou que as áreas da circulação que experimentam as maiores forças de cisalhamento são as mais propensas a desenvolver danos crônicos que eventualmente levam a doenças cardíacas.
O revestimento protetor do endotélio, o glicocálice, é estruturado (devido ao seu alto teor de sulfato) para criar grandes quantidades de gel (água líquida cristalina) ao seu redor e, assim, proteger o endotélio.
Além disso, uma camada fina na qual as células sanguíneas não podem entrar é conhecida por revestir diretamente as paredes até mesmo dos menores vasos sanguíneos. Da mesma forma, pesquisadores em 2000 observaram uma camada gelada de 0,4-0,5μm de espessura de água que reveste o endotélio de capilares saudáveis (os menores vasos sanguíneos do corpo), e esse revestimento exclui uma variedade de substâncias diferentes de atingir o endotélio.
Oficialmente, a proteína spike não pode afetar o endotélio porque não tem um receptor ACE2 funcional que a spike usa para entrar nele. Embora esse seja o caso, há muitas outras maneiras pelas quais a proteína spike ainda pode se ligar ao endotélio (por exemplo, a proteína spike ainda pode se ligar ao receptor, ela se liga diretamente à heparina no gliocálice e, se for produzida de dentro do endotélio pela spike, entrou efetivamente no endotélio).
Uma vez que autópsias de indivíduos suspeitos de terem morrido devido à mrna revelaram danos profundos à camada endotelial, isso fornece fortes evidências de que a proteína spike, de uma forma ou de outra, tem a capacidade de penetrar na barreira normalmente protetora.
Muitos suspeitaram que isso se deve em parte à carga altamente positiva que a proteína spike (e os componentes às vezes carregados positivamente das nanopartículas lipídicas) carrega, já que essas cargas positivas podem penetrar na barreira de água cristalina líquida que protege células como o endotélio.
Acredito que também pode ser devido à interrupção do potencial zeta da proteína spike, já que outros estados associados ao aumento da viscosidade do sangue (por exemplo, diabetes descontrolada) são conhecidos por danificar o endotélio e acelerar sua progressão para doenças cardíacas (além de aumentar o risco de diabetes).
Transições de fase
Embora a água líquida cristalina seja necessária para a vida, muito dela também pode ser problemática. Por exemplo, se toda a célula fosse coberta com água em gel, não seria possível que muitas coisas que precisavam entrar na célula entrassem.
Para resolver esse problema, existem áreas de carga positiva na membrana (por exemplo, Pollack argumenta que esta é uma razão chave para a colocação de íons metálicos dentro da membrana celular), que impedem a formação de água cristalina líquida sobre a área e, assim, criam uma passagem aberta para a água viajar.
Nota: embora sejamos ensinados a pensar na célula como um mosaico fluido de fosfolipídios com proteínas aqui e ali, aproximadamente 50% da membrana celular é tipicamente composta de proteínas (e às vezes pode ser ainda mais).
Na maioria dos casos, no entanto, o corpo depende de ser capaz de transformar a água entre seu estado de gel (líquido cristalino) e um estado de sol (água a granel contendo suspensões coloidais) conforme necessário.
No entanto, embora o reconhecimento da importância fundamental das mudanças de fase na biologia celular tenha aumentado gradualmente nos últimos 70 anos, a maioria do campo científico ainda está atrelada à hipótese do mosaico de fluidos, que acredita que a membrana celular sendo sempre um líquido é um dogma central da biologia celular estrutural.
Resumidamente, os géis (naturais e sintéticos) normalmente têm uma temperatura (seu ponto de transição) onde o gel passará por uma rápida transição de um estado para o outro (explicado mais adiante aqui).
Tanto quanto sei, em todos os casos, a temperatura mais quente corresponderá ao estado sol (H 2 O água com coloides em suspensão) e a mais fria corresponderá ao estado gel (H3O2). Além disso, o aumento da pressão também promoverá o estado do gel:
“A atividade biológica requer quase universalmente que as membranas estejam em seu estado fluido; em muitos casos, logo acima do ponto de transição”
Os géis em todo o corpo, por sua vez, são projetados para ter um ponto de transição de fase próximo à temperatura normal do corpo. Isso pode explicar por que a maioria dos mamíferos e aves mantém sua temperatura entre 35° – 42°C (95° – 107,6°F; a temperatura normal para humanos é de 37°C ou 98,6°F), um fato curioso dado o quanto seus corpos e ambientes diferem (por exemplo, um pequeno pássaro no deserto vs. uma baleia azul).
Além disso, essa temperatura se estende além dos animais de sangue quente. Um atum, quando ativo, mantém a temperatura de seu corpo em 30°C ou 86°F (o oceano é muito mais frio), enquanto muitos répteis mantêm sua temperatura corporal perto de 40°C ou 104°F (fazendo coisas como procurar o sol). Muitos insetos também não podem voar até que tenham se aquecido para uma faixa semelhante de temperatura (por exemplo, 38°C ou 100,4°F para abelhas).
Da mesma forma, o ponto de transição de fase é fortemente influenciado pelos materiais a partir dos quais o gel é construído (por exemplo, componentes da membrana celular). Animais de sangue frio, por sua vez, têm sido observados para alterar a composição de suas membranas celulares em resposta ao seu ambiente.
Em um experimento, Calotes versicolor (lagarto comum do jardim) foi aclimatado a 16°, 26° e 36°C (60,8°, 78,8° e 96,8°F) por um período de trinta dias. Os lagartos exibiram mudanças dramáticas na concentração dos principais fosfolipídios (componentes da membrana celular), como fosfatidilcolina, fosfatidiletanolamina, fosfatidilinositol, cardiolipina e esfingomielina.
A aclimatação à temperatura também tem sido observada para variar a composição lipídica dos microssomas do músculo da carpa juntamente com as células sanguíneas da carpa, e variação sazonal tem sido observada na composição lipídica do cérebro da truta arco-íris.
Uma vez que o gel é mantido perto de sua temperatura de transição, é necessário mudar exatamente onde essa temperatura de transição está para que a água esteja na fase apropriada para as necessidades fisiológicas atuais. Isso é feito principalmente dentro do corpo, alterando a composição eletrolítica da área, pois cátions fortes (íons carregados positivamente) destroem géis, enquanto ânions fortes (íons carregados negativamente) os reforçam.
Até onde eu posso dizer, os efeitos desses íons se correlacionam com os ânions de alta valência (carga) (-) que suportam mais efetivamente o potencial zeta fisiológico e aqueles cátions de alta valência (+) que mais efetivamente o destroem.
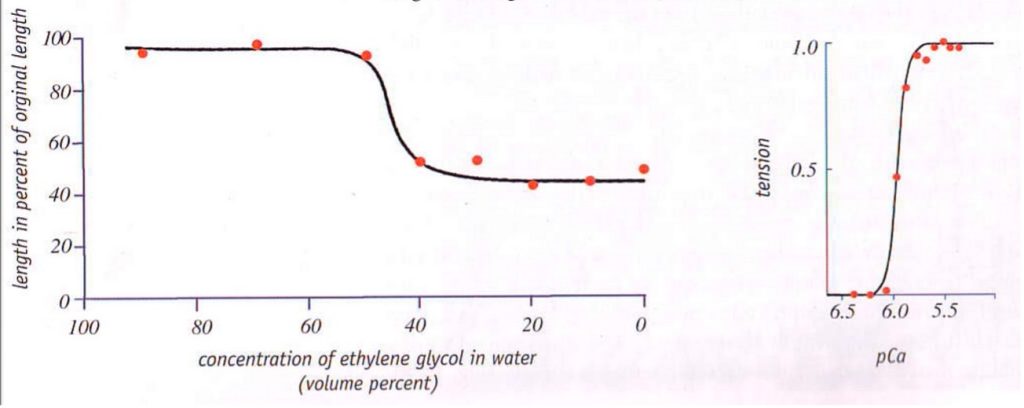
O principal elemento catiônico que o corpo usa para mudar a temperatura de transição a fim de destruir géis é o cálcio (Ca). Para ilustrar graficamente o efeito do cálcio (observe como um limiar crítico súbito de concentração de Ca é atingido):
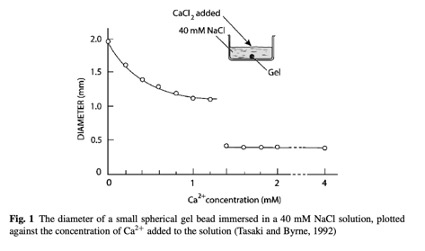
Esses resultados também mostraram como os cátions de valência mais alta, Ca2+, causam a formação de géis menores em relação aos cátions de baixa valência Na(1)+.
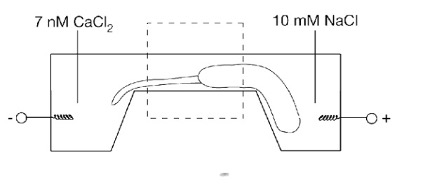
Depois de juntar tudo isso, você efetivamente obtém uma situação como essa dentro das células, o que permite que pequenas secreções de cálcio mudem sua água de um fluido para um estado de gel.
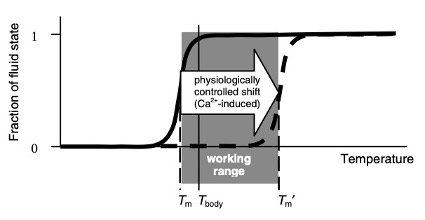
Por outro lado, se o corpo pode dispersar os géis, também precisa de uma maneira de recriá-los. O ânion forte primário (alta valência) que usa para construí-los são os fosfatos em trifosfato de adenosina (ATP).
Quando consideramos essa função alternativa do ATP, devemos considerar algumas das ideias propostas pela primeira vez por Ling. Eles sugerem que grande parte da energia obtida do ATP não surge da energia dentro de suas ligações sendo quebrada, mas sim da indução da água para formar uma estrutura cristalina líquida. Em essência, grande parte da energia gerada a partir do ATP surge, em última análise, da energia radiante que desencadeia essas polimerizações (géis).
Transições de fase são muito comuns na biologia celular, e embora o campo não coloque uma grande ênfase em sua importância, o argumento central de Pollack em seus dois primeiros livros é que as transições de fase são necessárias para explicar muitas funções da célula.
Géis de expansão
Um dos principais usos para géis dentro do corpo é expandir rapidamente géis e criar efeitos a partir dessa expansão. Por exemplo, as células frequentemente excretam pequenas esferas (conhecidas como vesículas) contendo componentes importantes, que ao sair da célula se expandem rapidamente.
Vasos isolados produtores de mucina, por exemplo, sofrem uma expansão de 600 vezes em 40 milissegundos.
Antes de sair da célula, os componentes internos da vesícula são ligados entre si por Ca2+ (lembre-se, cátions de valência mais alta, através de proteínas em gel de reticulação, limitarão a formação de gel, enquanto cátions monovalentes tendem a expandi-los).
Uma vez secretado da célula, o Ca2+ é removido e o gel hidrata-se rapidamente na solução de Na(1)+ que envolve cada célula [nota: os géis não se expandem rapidamente em água destilada, e foi observado que a formação de água líquida cristalina também é diminuída dentro da água deionizada].
Nota: Algumas vesículas utilizam fatores ambientais locais para mudar de fase. Por exemplo, a histamina é um cátion monovalente (carga de +1) em um pH neutro, mas se torna um cátion divalente (carga de +2) em um pH ácido.
Os pesquisadores conseguiram mostrar que a histamina em um ambiente ácido (devido ao seu alto estado de valência), causa encolhimento dos grânulos de mastócitos, enquanto a histamina em um ambiente neutro permite sua expansão.
Fisiologia Muscular
Um dos problemas mais comuns para os quais os pacientes procuram o médico são as dores musculoesqueléticas originadas por músculos excessivamente tensos. Por essa razão, analisei muitas abordagens diferentes para abordar essas questões.
Embora eu tenha encontrado alguns que eu sinto que são notavelmente eficazes, eu comumente acho que não há nenhum modelo existente dentro da fisiologia muscular que possa explicar por que muitos dos tratamentos que eu conheço realmente funcionam.
Um dos grandes problemas da medicina (e muitas vezes da ciência) é o que chamo de “armadilha mecanicista”. Resumidamente, isso significa que, se não há um mecanismo para explicar por que algo funciona, então é assumido como uma farsa e pensado para não funcionar de fato.
Acho que esse viés é bastante bobo, já que muitos dos modelos que usamos para explicar por que as drogas funcionam, na verdade, não fazem sentido.
Há muitos casos de um modelo previamente aceito para o mecanismo de uma droga que foi descartado e substituído por outro, e eu conheço muitos outros exemplos em que um modelo heterodoxo parece fornecer uma explicação muito melhor para o mecanismo de uma terapia.
Infelizmente, é quase impossível que o FDA (ou muitos médicos convencionalmente treinados) esteja aberto a considerar uma terapia para pesquisa, a menos que um mecanismo bioquímico para isso possa ser avançado.
Acredito que isso ocorre porque as drogas farmacêuticas, como são projetadas para atingir certas enzimas no corpo, se encaixam em nosso paradigma mecanicista rígido, enquanto terapias com um amplo espectro de defesa, que não podem ser ligadas a um receptor bioquímico específico, não.
Diante de tudo isso, fiquei muito feliz em descobrir através de Pollack que muitos aspectos do modelo existente para a fisiologia muscular não fazem sentido, e os resultados esperados do modelo não ocorrem quando experimentos em músculos são realizados:
No entanto, um modelo alternativo para a fisiologia muscular também existe. Antes de prosseguirmos, admito que sou um pouco tendencioso aqui porque esse modelo fornece um mecanismo para algumas das terapias que observei repetidamente tratar músculos hiperativos.
Quando ensino alunos e pacientes sobre músculos, explico-lhes que os músculos só podem contrair e não alongar. Sempre que uma parte do corpo “alonga” (por exemplo, estender a mão e tocar em algo à sua frente), isso ocorre porque um músculo que se contrai do outro lado de uma articulação fará com que a articulação se alongue.
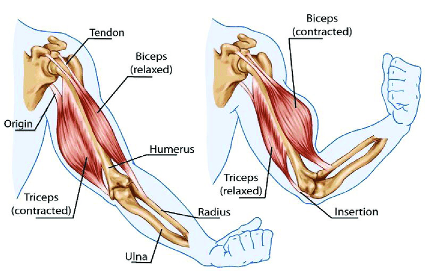
Para que o sistema funcione, ele sempre tem que ter um músculo que relaxe perfeitamente em sincronia com o músculo adversário que se contrai.
Infelizmente, em muitos casos, o músculo relaxante não consegue relaxar e, em vez disso, causa uma disfunção crônica ocorrer dentro da unidade muscular. Com Pollack, no entanto, descobri que realmente havia feito uma suposição errônea por anos.
O modelo de Pollack foi baseado na observação de que os músculos relaxados contêm uma grande quantidade de água estruturada, enquanto os músculos contraídos contêm principalmente água que não está em um estado de gel semissólido.
Uma vez que se sabe que o disparo muscular segue o sinal nervoso que direciona os íons cálcio (Ca2+) para entrar no músculo, isso argumenta para um modelo distintamente diferente para explicar a fisiologia muscular.
Por outro lado, também se sabe (a partir do experimento descrito anteriormente na seção de gradiente) que, enquanto as células musculares que foram cortadas podem reter seu gradiente de potássio para sódio enquanto em uma solução salina, as partes que carecem de ATP são incapazes de fazê-lo.
Isso sugere que o ATP é necessário para que o estado líquido cristalino da água esteja presente, pois exclui a entrada de íons sódio nas células musculares. Da mesma forma, sabe-se também que, sem ATP, os músculos permanecerão presos na forma contraída e serão incapazes de se alongar, algo que ocorre logo após a morte (denominado “rigor mortis“).
Pollack também observou que as mudanças musculares em resposta a fatores ambientais seguiram um limiar crítico sendo ultrapassado. Isso parecia sugerir a Pollack que uma mudança de fase estava ocorrendo dentro dos músculos.
Então, qual é o modelo de Pollack? Os músculos são projetados para formar grandes géis ao redor deles e, à medida que a água cristalina líquida dentro deles se expande, ela estica as proteínas não enroladas nas fibras em hélices (até que as ligações cruzadas no músculo impeçam o alongamento adicional).
Quando o músculo precisa se contrair, os íons de cálcio entram no músculo e eliminam os géis, e as proteínas que não estão mais alongadas elasticamente voltam à sua configuração original sem bobina. Para que o gel se reconstrua, é necessário que o ATP quebre as ligações cruzadas criadas pelo cálcio para desencadear uma mudança de fase que reconstrói o gel.
Assim, a força de um músculo é um produto da energia potencial criada pelo armazenamento de água líquida cristalina de energia radiante sendo liberada. Ou, dito de outra forma, a força dos músculos na verdade está em sua capacidade de alongar espontaneamente, algo que eu havia assumido anteriormente que eles não poderiam fazer. Essa mecânica física de todo esse processo ajuda a explicar muitas das minhas observações sobre os músculos:
O grande cisalhamento mecânico faz com que a matriz do citoesqueleto [outra estrutura-chave que se expande e se contrai em resposta a mudanças de fase] fluidifique, um fenômeno semelhante ao rejuvenescimento físico em SGM’s [certos materiais sintéticos].
Essa fluidização é seguida por uma recuperação lenta e livre de escala das propriedades mecânicas, um fenômeno semelhante ao envelhecimento físico em SGMs. Surpreendentemente, em resposta a um estiramento transitório, o citoesqueleto fluidiza em um padrão que é universal para diferentes tipos celulares.
Esse achado implica mecanismos mediados não tanto por vias de sinalização específicas, como se costuma supor, mas sim – como explicamos a seguir – por ações não específicas de forças físicas.
Muitas outras estruturas também têm a capacidade de seus géis internos causarem expansão e contração, mas ao contrário dos músculos esqueléticos, na maioria dos casos, essa expansão e contração não é restringida pelas ligações cruzadas entre as proteínas musculares.
Músculos lisos, bem como anéis contráteis, géis sintéticos de actina (actina é uma das proteínas contráteis mais comuns no corpo) e outras organelas com poucas ligações cruzadas covalentes e estruturas semi-aleatórias, podem assim encurtar maciçamente.
Por fim, também foi observado que alterar as concentrações circundantes de salinidade ao redor de outras fibras (por exemplo, colágeno) pode fazer com que elas se expandam e se contraiam a ponto de gerar uma força que produz contração como uma fibra muscular.
Conclusão
Houve três questões centrais em que pensei ao escrever esta série. A primeira é que muitos sistemas médicos tradicionais acreditam que existe uma força expansiva inerente em todo o corpo que desempenha um papel fundamental no fornecimento de sua vitalidade, enquanto quando está comprometida, uma variedade de doenças se instala.
Acredito que seja esse o caso, pois na prática, torna-se possível reconhecer onde essa expansão foi substituída por um espaço compactado e, ao longo dos séculos, algumas abordagens realmente intrigantes foram desenvolvidas que parecem abordar esse problema e restaurar a vitalidade de um paciente. Infelizmente, não existe um modelo que explique de onde vem essa força expansiva.
O segundo é qual é exatamente a relação entre o potencial zeta e a presença de água líquida cristalina no corpo (passei mais tempo do que posso contar tentando entender isso). Como mencionado anteriormente, praticamente todos os sistemas coloidais naturais dependem de carga negativa mútua para dispersão.
Da mesma forma, a água líquida cristalina é tipicamente carregada negativamente e formada em superfícies carregadas negativamente. A razão pela qual o cálcio (que prejudica o potencial zeta) condensa tão eficazmente as matrizes proteicas que dão origem aos géis (destruindo assim os géis), deve-se às proteínas que contêm cargas negativas às quais o cálcio pode efetivamente ligar-se.
O terceiro é onde o movimento surge de dentro do corpo, já que o comprometimento de qualquer tipo de circulação (existem muitas diferentes dentro do corpo) é uma das causas mais comuns de doenças.
Neste artigo, tentei ilustrar as consequências estruturais da água líquida cristalina dentro do corpo, e espero ter respondido à primeira pergunta. No resto desta série, explorarei essas duas próximas questões juntamente com as outras maneiras pelas quais a água estruturada pode transmitir informações e gerar energia dentro do corpo (por exemplo, com os nervos), e então concluirei com as abordagens que conheço para aumentar a geração dessa fase da água dentro do corpo.
Espero que você esteja achando esta série interessante e que, mesmo que você não consiga entender tudo (estou tentando encontrar um bom equilíbrio entre tornar o assunto compreensível e ter as informações necessárias para partes mais céticas), espero que ainda haja algumas dicas úteis que você possa levar para casa a partir disso.
Como isso é um pouco técnico e isso leva muito tempo para ser escrito, vou voltar um pouco ao assunto, para que você possa digerir as duas primeiras partes desta série. Embora eu tenha feito o meu melhor (reli tudo aqui e mais para esta série) como não sou Pollack, é muito provável que eu esteja interpretando mal alguns de seus trabalhos.
Se você está familiarizado com qualquer um desses assuntos, eu gostaria muito de saber se você acredita que eu cometi algum erro significativo aqui.



